1. Bucher P. Weight matrix descriptions of four eukaryotic RNA polymerase II promoter elements derived from 502 unrelated promoter sequences. J Mol Biol. 1990;212(4):563-578. 10.1016/0022 2836(90)90223-9. DOI: 10.1016/00222836(90)90223-9
2. Burley S.K., Roeder R.G. Biochemistry and structural biology of transcription factor IID (TFIID). Annu Rev Biochem. 1996;65:769-799. DOI: 10.1146/annurev.bi.65.070196.004005
3. Burow M., Atwell S., Francisco M., Kerwin R.E., Halkier B.A., Kliebenstein D.J. The glucosinolate biosynthetic gene AOP2 mediates feedback regulation of jasmonic acid signaling in Arabidopsis. Mol Plant. 2015;8(8):1201-1212. DOI: 10.1016/j.molp.2015.03.001
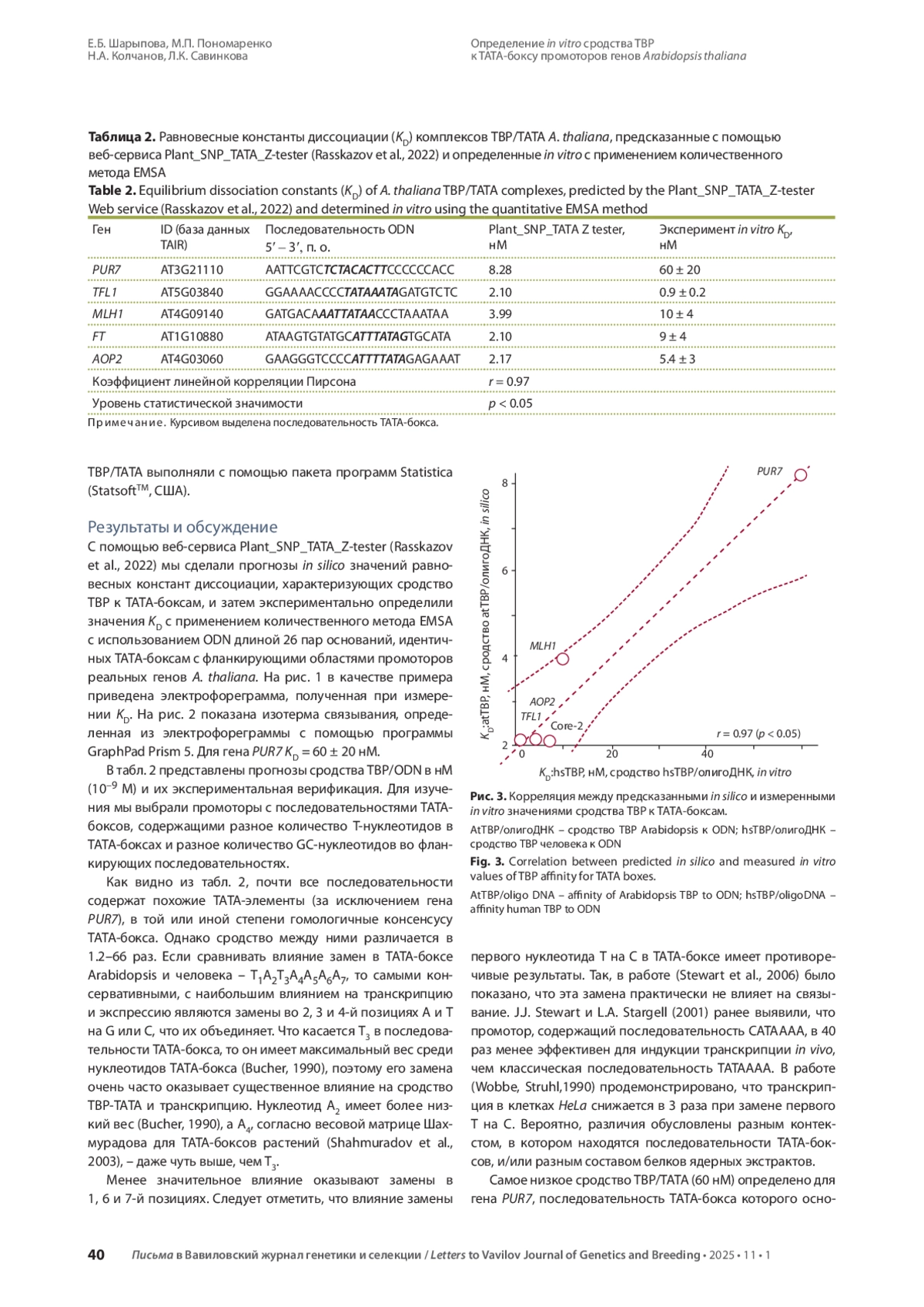
4. Cerise M., da Silveira Falavigna V., Rodríguez-Maroto G., Signol A., Severing E., Gao H., van Driel A., Vincent C., Wilkens S., Iacobini F.R., Formosa-Jordan P., Pajoro A., Coupland G. Two modes of gene regulation by TFL1 mediate its dual function in flowering time and shoot determinacy of Arabidopsis. Development. 2023;150(23):dev202089. DOI: 10.1242/dev.202089
5. Corbesier L., Vincent C., Jang S., Fornara F., Fan Q., Searle I., Giakountis A., Farrona S., Gissot L., Turnbull C., Coupland G. FT protein movement contributes to long-distance signaling in floral induction of Arabidopsis. Science. 2007;316(5827):1030-1033. 10.1126/ science.1141752. DOI: 10.1126/science.1141752
6. Das B. Glucosinolate biosynthesis: role of MAM synthase and its perspectives. Biosci Rep. 2021;41(10):BSR20211634. 10.1042/ BSR20211634. DOI: 10.1042/BSR20211634
7. Das S., Bansal M. Variation of gene expression in plants is influenced by gene architecture and structural properties of promoters. PLoS One. 2019;14(3):e0212678. DOI: 10.1371/journal.pone.0212678 EDN: NGZSHX
8. Dikstein R. The unexpected traits associated with core promoter elements. Transcription. 2011;2(5):201-206. DOI: 10.4161/trns.2.5.17271
9. Dion E., Li L., Jean M., Belzile F. An Arabidopsis MLH1 mutant exhibits reproductive defects and reveals a dual role for this gene in mitotic recombination. Plant J. 2007;51(3):431-440. 10.1111/ j.1365-313X.2007.03145.x. DOI: 10.1111/j.1365-313X.2007.03145.x
10. Drachkova I.A., Ponomarenko P.M., Arshinova T.V., Ponomarenko M.P., Suslov V.V., Savinkova L.K., Kolchanov N.A. In vitro examining the existing prognoses how TBP binds to TATA with SNP associated with human diseases. Health. 2011;3(9):577-583. 10.4236/ health.2011.39099. DOI: 10.4236/health.2011.39099
11. Faiger H., Ivanchenko M., Cohen I., Haran T.E. TBP flanking sequences: asymmetry of binding, long-range effects and consensus sequences. Nucleic Acids Res. 2006;34:104-119. DOI: 10.1093/nar/gkj414 EDN: ITYGVV
12. Garcia-Hernandez M., Berardini T.Z., Chen G., Crist D., Doyle A., Huala E., Knee E., Lambrecht M., Miller N., Mueller L.A., Mundodi S., Reiser L., Rhee S.Y., Scholl R., Tacklind J., Weems D.C., Wu Y., Xu I., Yoo D., Yoon J., Zhang P. TAIR: a resource for integrated Arabidopsis data. Funct Integr Genomics. 2002;2(6):239-253. DOI: 10.1007/s10142-002-0077-z
13. Goretti D., Silvestre M., Collani S., Langenecker T., Méndez C., Madueño F., Schmid M. TERMINAL FLOWER1 functions as a mobile transcriptional cofactor in the shoot apical meristem. Plant Physiol. 2020;182(4):2081-2095. DOI: 10.1104/pp.19.00867 EDN: VQXPTM
14. Hampsey M. Molecular genetics of the RNA polymerase II general transcriptional machinery. Microbiol Mol Biol Rev. 1998;62:465-503. DOI: 10.1128/MMBR.62.2.465-503.1998
15. Holdsworth M.J., Grierson C., Schuch W., Bevan M. DNA-binding properties of cloned TATA-binding protein from potato tubers. Plant Mol Biol. 1992;19:455-464. DOI: 10.1007/BF00023393
16. Iwataki N., Hoya A., Yamazaki K. Restoration of TATA-dependent transcription in a heat-inactivated extract of tobacco nuclei by recombinant TATA-binding protein (TBP) tobacco. Plant Mol Biol. 1997;34:69-79. :1005759521285. DOI: 10.1023/A
17. Juven-Gershona T., Kadonaga J.T. Regulation of gene expression via the core promoter and the basal transcriptional machinery. Dev Biol. 2010;339(2):225-229. DOI: 10.1016/j.ydbio.2009.08.009 EDN: NYZCCN
18. Kadonaga J.T. Perspectives on the RNA polymerase II core promoter. Wiley Interdiscip Rev Dev Biol. 2012;1(1):40-51. DOI: 10.1002/wdev.21
19. Lv X., Zeng X., Hu H., Chen L., Zhang F., Liu R., Liu Y., Zhou X., Wang C., Wu Z. Structural insights into the multivalent binding of the Arabidopsis FLOWERING LOCUS T promoter by the CO-NF-Y master transcription factor complex. Plant Cell. 2021;33(4):1182-1195. DOI: 10.1093/plcell/koab016 EDN: LLCQGW
20. Marks R.A., Hotaling S., Frandsen P.B., VanBuren R. Representation and participation across 20 years of plant genome sequencing. Nat Plants. 2021;7(12):1571-1578. doi 0.1038/s41477-021-01031-8.
21. Mogno I., Vallania F., Mitra R.D., Cohen B.A. TATA is a modular component of synthetic promoters. Genome Res. 2010;20(10):1391-1397. DOI: 10.1101/gr.106732.110
22. Morey C., Mookherjee S., Rajasekaran G., Bansal M. DNA free energybased promoter prediction and comparative analysis of Arabidopsis and rice genomes. Plant Physiol. 2011;156(3):1300-1315. DOI: 10.1104/p.p.110.167809
23. Mukumoto F., Hirose S., Imaseki H., Yamazaki K.-I. DNA sequence requirement of a TATA element-binding protein from Arabidopsis for transcription in vitro. Plant Mol Biol. 1993;23:995-1003. DOI: 10.1007/BF00021814
24. Patikoglou G.A., Kim J.L., Sun L., Yang S.H., Kodadek T., Burley S.K. TATA element recognition by the TATA box-binding protein has been conserved throughout evolution. Genes Dev. 1999;13(24):3217-3230. DOI: 10.1101/gad.13.24.3217
25. Ponomarenko M., Rasskazov D., Arkova O., Ponomarenko P., Suslov V., Savinkova L., Kolchanov N. How to use SNP_TATA_Comparator to find a significant change in gene expression caused by the regulatory SNP of this gene’s promoter via a change in affinity of the TATAbinding protein for this promoter. Biomed Res Int. 2015;2015:359835. DOI: 10.1155/2015/359835
26. Porto M.S., Pinheiro M.P., Batista V.G., dos Santos R.C., Filho Pde A., de Lima L.M. Plant promoters: an approach of structure and function. Mol Biotechnol. 2014;56(1):38-49. DOI: 10.1007/s12033-013-9713-1 EDN: SPYGUL
27. Pugh B.F. Purification of the human TATA-binding protein, TBP. In: Tymms M.J. (Ed.) In Vitro Transcription and Translation Protocols. (Ser. Methods in Molecular Biology. Vol. 37). Totowa, NJ: Humana Press Inc, 1995;359-367. :359. DOI: 10.1385/0-89603-288-4
28. Rasskazov D., Chadaeva I., Sharypova E., Zolotareva K., Khandaev B., Ponomarenko P., Podkolodnyy N., Tverdokhleb N., Vishnevsky O., Bogomolov A., Podkolodnaya O., Savinkova L., Zemlyanskaya E., Golubyatnikov V., Kolchanov N., Ponomarenko M. Plant_SNP_TATA_Ztester: a Web service that unequivocally estimates the impact of proximal promoter mutations on plant gene expressions. Int J Mol Sci. 2022;23(15):8684. DOI: 10.3390/ijms23158684
29. Rhee H.S., Pugh B.F. Genome-wide structure and organization of eukaryotic pre-initiation complexes. Nature. 2012;483(7389):295-301. DOI: 10.1038/nature10799
30. Sandelin A., Carninci P., Lenhard B., Ponjavic J., Hayashizaki Y., Hume D.A. Mammalian RNA polymerase II core promoters: insights from genomewide studies. Nat Rev Genet. 2007;8:424-436. DOI: 10.1038/nrg2026 EDN: MMEQWN
31. Savinkova L., Drachkova I., Arshinova T., Ponomarenko P., Ponomarenko M., Kolchanov N. An experimental verification of the predicted effects of promoter TATA-box polymorphisms associated with human diseases on interactions between the TATA boxes and TATAbinding protein. PLoS One. 2013;8(2):e54626. DOI: 10.1371/journal.pone.0054626 EDN: RFDWST
32. Senecoff J.F., McKinney E.C., Meagher R.B. De novo purine synthesis in Arabidopsis thaliana. II. The PUR7 gene encoding 5’-phosphoribosyl-4-(N-succinocarboxamide)-5-aminoimidazole synthetase is expressed in rapidly dividing tissues. Plant Physiol. 1996;112(3):905-917. DOI: 10.1104/pp.112.3.905
33. Shahmuradov I.A., Gammerman A.J., Hancock J.M., Bramley P.M., Solovyev V.V. PlantProm: a database of plant promoter sequences. Nucleic Acids Res. 2003;31(1):114-117. DOI: 10.1093/nar/gkg041 EDN: MDFRBL
34. Shannon S., Meeks-Wagner D.R. Genetic interactions that regulate inflorescence development in Arabidopsis. Plant Cell. 1993;5(6):639-655. DOI: 10.1105/tpc.5.6.639
35. Song Y.H., Shim J.S., Kinmonth-Schultz H.A., Imaizumi T. Photoperiodic flowering: time measurement mechanisms in leaves. Annu Rev Plant Biol. 2014;66:441-464. DOI: 10.1146/annurev-arplant-043014-115555
36. Stewart J.J., Stargell L.A. The stability of the TFIIA-TBP-DNA complex dependent on the sequence of the TATAAA element. Biol Chem. 2001;276:30078-30084. DOI: 10.1074/jbc.M105276200
37. Stewart J.J., Fischback J.A., Chen X., Stargell L.A. Non optimal TATA element exibits diverse mechanistic consequences. J Biol Chem. 2006;281:22665-22673. DOI: 10.1074/jbc.M603237200
38. Wobbe C.R., Struhl K. Yeast and human TATA-binding proteins have nearly identical DNA sequence requirements for transcription in vitro. Mol Cell Biol. 1990;10:3859-3867. DOI: 10.1128/MCB.10.8.3859
39. Yamaguchi Y., Itoh Y., Takeda Y., Yamazaki K. TATA sequence requirements for the initiation of transcription for an RNA polymerase II in vitro transcription system from Nicotiana tabacum. Plant Mol Biol. 1998;38(6):1247-1252. :1006056128129. DOI: 10.1023/a